HOX gene collinearity- comparison of models
Dedicated to the memory of Nigel Holder-
an open-minded scientist and friend
Introductory note
I was fascinated by Hox gene collinearity from the first moment I came across this multiscale phenomenon. As a Physicist, I instantly thought that somehow a physical mechanism must be at the origin of this enigma. My conviction was reinforced every time a facet of the phenomenon was illuminated by new experimental evidence. I elaborated my idea step by step and I formulated a model which explains satisfactorily, to my taste, almost all existing data. In working out my model, the biophysical model, I did not have the opportunity to discuss in depth this issue and related problems. In my home Institute at ‘Demokritos’, my colleagues and friends are involved in research in quite different areas. Colleagues abroad are inclined to endorse more conventional descriptions – molecular and gene mechanisms that have been proposed. As a result, my proposition has been formulated in vacuum and I accept full responsibility for it. From the accumulating evidence I feel confident that my model is correct. However, one cannot exclude a sudden knock-down. In the endless effort of model verification one can only establish the compatibility between experiment and theory. As Karl Popper convincingly argued, one can never prove experimentally a theory but one can surely experimentally disprove it.
Having Popper’s aphorism in mind, I propose below two experiments for which the different models provide divergent predictions. Thus, if these experiments are ever performed, it will be possible to distinguish which model (if any) is more appropriate to explain the collinearity of Hox genes.
In the following exposition, the existing up to date results are briefly presented together with the proposed models for their explanation. I try to give an objective account of the different views but, admittedly, I do not hide my preference for the biophysical model. Therefore, I am eagerly expecting any kind of comments (positive or negative) from the readership of this blog. I only expect the objective readers to start doubting for the validity of the trendy dogmas on Hox gene collinearity when there are indications that such dogmas are inadequate or wrong.
English is not my mother language therefore my writings contain several syntax and grammatical errors. Nevertheless I prefer to leave the text with its linguistic mistakes since this informal style suits me better to convey directly my ideas.
------
Vertebrate data (older and new)
E.B.Lewis was the first who noticed the strange feature of collinearity for some homeotic genes in the Drosophila embryo, the so-called Hox genes [1].
At the nuclear (microscopic) level, these genes are clustered in a particular chromosome. The cluster of the ordered genes Hox1, Hox2, Hox3, … is located along the 3’ to 5’ direction on the chromosome. At the multicellular (macroscopic) level of the vertebrate embryo, the genes of the Hox cluster are activated along the anterior-posterior axis of the embryo in a partially overlapping manner (Fig.1): the anterior boundary of expression of a Hox gene is shifted posteriorily compared to the anterior boundary of expression of the precedent Hox gene in the sequence Hox1, Hox2, Hox3,…(spatial collinearity) : The microscopic axis 3’ to 5’ is collinear with the macroscopic axis anterior-posterior. The vertebrates possess four paralogous Hox clusters (HoxA, HoxB, HoxC and HoxD) each one positioned on a different chromosome [2]. This strange property of Hox gene expressions indicates that a profound correlation is at work between the macroscopic scale of the embryo (of the order of 1 mm) and the microscopic scale of the chromatin domain (of about 500 nm). Typically the scale difference extends in 3 orders of magnitude [3]. This multiscale correlation is an organization characteristic of systems biology [4].
Besides spatial collinearity, it was found that Hox1, Hox2, Hox3,…of a Hox cluster are activated sequentially in time following the same order (Fig.1): first Hox1, second Hox2,… (temporal collinearity) [5]. Furthermore, a third kind of collinearity was established (Fig.1): when at a given position on the anterior-posterior axis several Hox genes are co-activated, the expression of the most posterior gene in the cluster is stronger than the other gene expressions (quantitative collinearity) [6].

Schematic representation of the Hox gene expressions
The anterior boundary of Hox1 expression is located more anteriorily than the anterior boundary of Hox2 expression etc (spatial collinearity). The starting time (t1) of Hox1 expression is earlier than starting time (t2) of Hox2 expression etc (temporal collinearity). At a given position along the anterior-posterior axis (dashed line) the expression of Hox3 is stronger than the expression of Hox2 and Hox1 (quantitative collinearity).
The above impressive and well-known facts are described in several reviews [2,7]. The last decade some genetic engineering methods were developed (almost exclusively in Duboule’s Lab) which make possible the accurate intervention in the Hoxd locus and, as a result, transgenic mice are created with deleted or duplicated regions of the Hoxd cluster [8,9]. In other experiments Hoxb1 is transposed in the Hoxd cluster [10]. The produced transgenic expressions are compared to the wild type expressions in the mice limb buds and the trunk [8,9]. This comparison is very interesting since it illuminates several facets of the mechanism responsible for the collinearity of Hox genes.
Some months ago Tschopp and Duboule separated the centromeric neighborhood of the Hoxd cluster from the cluster itself, by engineering a large inversion of this centromeric neighborhood. The aim was to test the regulatory ‘landscape effect’ over the activity of the Hoxd cluster [11].
Collinearity models
In the following I will describe briefly two distinct models formulated in order to explain the Hox gene collinearity.
1. The two-phases model (T-PM) was put forward by Duboule and co-workers and the details are extensively presented in references [8,9]. It is a molecular model that functions at the early phase of mouse development (up to about stage E9.5) and the late phase (up to about E12.5). In the early phase, gene activation is regulated sequentially in time from the telomeric side of the Hoxd cluster. This positive activation is balanced by a repressive centromeric influence. The two influences combine and produce a sequential expression pattern in the direction 3’ to 5’.
2. The details of the biophysical model are presented elsewhere [ 3,12,13]. Here I would like to stress the recent illuminating evidence from the extensive work in the laboratory of P. R. Cook on the dynamics of gene activation [14]. It is generally believed that the movement of the polymerase (active and inactive) toward DNA causes gene activation. In contrast, Cook and his co-workers observed clearly that it is the DNA that moves toward the transcription factories where the polymerases are immobilized. This picture fits well to the biophysical model hypothesis: physical forces decondense and pull the chromatin fiber from inside the chromosome territory toward the transcription factories located in the interchromosome domain (Fig.2). An elastic spring expansion is the mechanical analogue of this chromatin decondensation and extrusion. When a gene moves away from the ‘factory’ the intensity of its activation drops sharply [15]. This observation offers a natural explanation of quantitative collinearity [13]. The biophysical model successfully describes almost all accumulated findings for the primary anterior-posterior axis and the secondary limb bud axis of vertebrates. The details are found in the publications [3,12,13]. However, these papers appeared before the very recent observations of Tschopp and Duboule [11]. I undertake here the challenge to account for these experiments of centromeric inversion by applying the biophysical model. Indeed, the biophysical model can satisfactorily explain the results (see below).

Mechanical analogue of Hox cluster decodensation and extrusion
a) Before activation the Hox cluster is condensed inside the chromatin territory
(CT)-(left). Mechanical analogue: an uncharged elastic spring fixed at its left end
(right).
b) The cluster is slightly decondenced and Hox1 is extruded in the
interchromosome domain (ICD) in the area of the transcription factory (TF -red
disc) (left). A small force F1 is applied at the loose end and expands slightly the
spring (right).
c) The cluster is further decondenced and the extruded Hox2 is located in the
transcription factory area while Hox1 moves away from TF (left). A bigger force
F2 expands further the spring (right).
Pros and Cons for the two models
The ‘two-phases model’ is based on well studied mechanisms involving enhancers, inhibitors, promoters and other molecules that regulate the genetic activity. Without excluding these important processes, the ‘biophysical model’ proposes an underlying mechanism that triggers where and when this molecular machinery is activated. Comparing the two models I would like to point out some differences.
1. The two-phases model extends to both early and late developmental phases aiming to explain the observed phenomena during all these stages. In contrast, the biophysical model is not so ambitious limiting its range to the early phase only when the responsible mechanism is relatively simpler.
2. As already mentioned, Hox gene collinearity is fundamentally a multiscale phenomenon where multicellular (macroscopic) and subcellular dimensions (microscopic) are inherently interconnected. This phenomenon is a characteristic example of systems biology with the multiscale organization requiring, besides the molecular, a multidisciplinary (physical and mathematical) treatment [4].
The biophysical model establishes such a multiscale interrelation: a spatial and temporal signal in every cell of the multicellular tissue is transduced to the genetic subcellular domain [3]. At this microscopic level, physical forces are generated which cause differential Hox gene activation. These microscale forces inherently contain the ‘positional-and-time information’ from the macroscale domain. Subsequently, the genetic activation is collectively incorporated in the multicellular level causing the characteristic expression patterns in space and time (Fig.3). The transition from the macroscopic to the microscopic and back again to the macroscopic scale is achieved by feedback loops which are indispensable in the multiscale organization of systems biology [4].
The two-phases model functions at the DNA (microscopic) level. The spatial demarcation of the Hox gene expressions at the tissue level is an observed (macroscopic) result without any causal relation or feedback from the microscopic scale of the model. The phenomena at the two different scales are schematically juxtaposed with no internal connection between them.
3. Quantitative collinearitry is naturally explained by the biophysical model: hox genes approach the transcription factory one after the other and subsequently they move away from it (Fig.2). In this process the closer a gene comes to the polymerase the stronger its expression [14,15]. Quite recent evidence supports the view of chromatin moving toward the immobile polymerase [15].
The two-phases model cannot reproduce the sequential intensity of hox gene expressions. In order to do this one has to make additional ad hoc assumptions.

Hox1 expression as a result from a multiscale action
Top: A macroscopic concentration gradient where the threshold T1 determines the
area for Hox1 expression along the anterior-posterior axis.
Center (right): The ‘positional information’ together with the temporal signal are
transduced inside the (microscopic) nucleus. A pulling force F1 is generated.
Bottom: Collective gene activation leads to the macroscopic Hox1 expression.
4. Several experimental findings are unexplained by the two-phases model but they are well reproduced by the biophysical model. For instance:
a) after the transposition of Hoxb1 in the Hoxd cluster, the cluster decondences but does not loop out [10]. This ‘unexpected’ result is naturally explained by the biophysical model [13].
b) For posterior deletions in the limb bud, according to the two-phases model, ‘unexpected’ redistributions of probe hox genes were observed [8]. These redistributions (posteriorizations) are predicted by the biophysical model [12].
c) For anterior deletions in the trunk, according to the two-phases model, ‘it is impossible to anticipate’ an up-regulation of the mutant hox gene expressions [9]. In contrast the biophysical model predicts an anterior extension of these expressions [12,13].
Doubt for the ‘landscape effect’ at the early phase
In the recent paper of Tschopp and Duboule the following genetic engineering experiment is described [11]: a large centromeric region neighboring the posterior end of the Hoxd cluster is inverted (Fig.4). For this inversion and among other observations, the transgenic expressions of Hoxd13-Hoxd10 at the early stages are compared to the wild type expressions. They find that the transgenic expressions are premature and spatially more extensive. They put forward the hypothesis that in this large centromeric region some smaller regions are contained which produce an inhibitory effect on the Hoxd cluster. Candidate regions causing this inhibitory effect have been localized centromeric to Evx2. Such regions lie e.g. between the Rel3 and Rel2 breakpoints [11, 16]. Following the two-phases model, Tschopp and Duboule [11] conclude that the above inversion relocates the regulatory centromeric region far away from the cluster so that its inhibitory influence on the cluster fades out. As a result, the centromeric ‘landscape effect’ is removed and the remaining positive telomeric influence causes the observed premature up-regulation of the posterior Hoxd expressions.
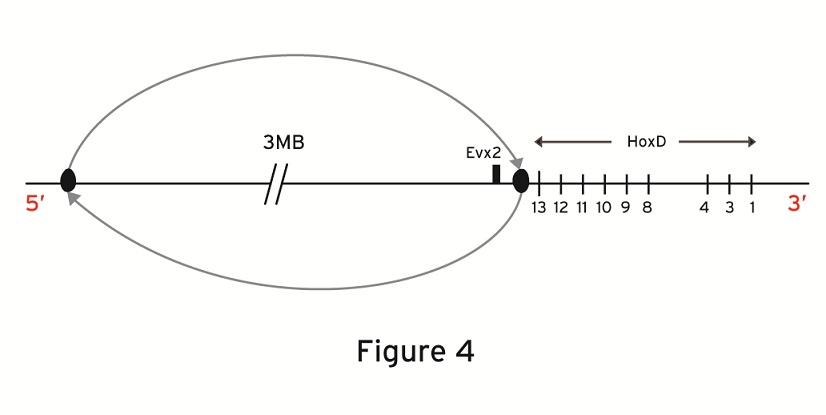
Large inversion centromeric to the HoxD cluster. Evx2 is included in the genetic
inversion.
The biophysical model proposes a quite different explanation of the above transgenic expressions (Fig.2): the mechanical analogue of the Hoxd cluster is the expanding elastic spring with a loose telomeric end (3’) and a fixed centromeric end located between the last gene of the cluster (Hoxd13) and Evx2 [13]. When the fixed end is removed the spring becomes loose at its both ends and it can slide and decondense when a smaller than normal force is applied to the telomeric end. The smaller force is related to a premature and more anterior than normal gene activation at the early stages [12,13]. This argumentation was used to explain the mutant expressions of Hoxd10 and Hoxd9 [13] when the engineered inversion included the posterior subcluster up to Hoxd11 [9].
The same explanation holds when the inversion including Evx2 starts just after Hoxd13 ( Fig.4). In this case, the fixed posterior end is cut-off and the elastic spring becomes loose at its both ends. Again, an extended activation (anteriorization) of Hoxd13-Hoxd10 is expected to occur prematurely. This is exactly what Tschopp and Duboule observe [ref.11: Fig 3]. If the biophysical model explanation is correct, the experimental evidence does not necessarily lead to ‘a landscape effect’ for the early stages. On the contrary, it reinforces the picture of the Hox cluster behaving like an elastic spring. At later stages however, secondary and restoring mechanisms get involved and such a ‘landscape effect’ may occur.
Proposal for two model-killer experiments
I presented above two distinct explanations for the experiment where a large centromeric inversion was genetically engineered. Is it possible to conclude which explanation is correct? In the following I propose two experiments which will help to decide.
A) Experimental design: in the proposed centromeric inversion shown in Fig.5, the inverted region is almost the same with the reported inversion of Fig.4. The only difference is that in the new inversion the small region between Hoxd13 and Evx2 is unaffected.
1. According to the two-phases model, this small region cannot significantly influence the ‘landscape effect’ which is due to the much bigger centromeric area that is inverted. Therefore it is expected that, at the early stages, the transgenic expressions of Hoxd13-Hoxd10 for both inversions (Fig.4 and Fig.5) should be almost the same: premature and overextended compared to the wild type expressions.
2. According to the biophysical model, the Hox cluster behaves like an elastic spring whose fixed posterior end lies in the small region between Hoxd13 and Evx2. As long as the fixed end of the spring remains in place the cluster behaves normally.

Large inversion centromeric to the HoxD cluster. Evx2 is not included in the genetic
inversion.
Therefore, for the inversion of Fig.5 during the early stages, the expressions of Hoxd13-Hoxd10 should be very similar to the wild type expressions.
Summarizing for the centromeric inversion of Fig.5, at the early phase, the two models predict quite different transgenic expressions of Hoxd13-Hoxd10: whereas the two-phases model predicts abnormal expressions (premature and up-regulated) - the biophysical model predicts normal (wild type) expressions. I think it should be of interest if this experiment were ever performed. If the prediction of one of the two models is confirmed, this will lead automatically to the refutation of the other model. There is still the possibility that the experimental result will differ substantially from both model predictions. In this case both models will be in trouble.
B) Another direct experiment could be the deletion of the small region between Hoxd13 and Evx2: according to the two-phases model this small deletion should not affect substantially the Hoxd expressions, so the mutant Hoxd expressions should be similar in space and time to the wild type ones. By contrast, according to the biophysical model arguments presented above, these mutant Hoxd expressions should be prematurely anteriorized in the early phase. The Kondo and Duboule experiment and its ‘unexpected’ findings (Cell 97, 407, (1999)) indirectly confirm the biophysical model prediction (see my blog below)!
The Evx2-Hoxd13 intergenic region of 8 kb was examined in detail and it was found that it plays a role as a boundary element with a differential spatial and temporal activity [17]. From this analysis it is clear that the deletion of this intergenic region is not expected to cause an effect on the Hoxd cluster comparable to the biophysical model prediction –premature and anteriorily ectopic Hoxd expressions in the early stages.
In case the biophysical model is demolished by the above experiments, my decision is not to write ever again a word on Hox gene collinearity!! However, if the experimental findings agree with the biophysical model predictions, it should be worth formulating some more detailed models or alternatives e.g.: a) What is a realistic distribution of the positive and negative electric charges and, taking into account their relative distances, what is the consequent accurate Coulomb force F? b) What is the degree of reversibility of the chromatin expansion -how elastic is the ‘spring’. c) Besides the Coulomb forces are there some other forces that can pull the chromatin fiber?
I am told that, although feasible, it is very difficult to carry out the proposed experiments because the genetic engineering techniques involved are very complicated and, furthermore, they are time consuming and expensive. The ideal place would be Duboule’s lab but this does not seem feasible.
References
1. Lewis, E.B. 1978. A gene complex controlling segmentation in Drosophila. Nature 276, 565-570.
2. Nolte, C. & Krumlauf, R. 2007. Expression of Hox genes in the nervous system of Vertebrates. In HOX gene expression. (Ed. S. Papageorgiou), pp. 14-41. Landes Bioscience & Springer , New York
3. Papageorgiou, S. 2006. Pulling forces acting on Hox gene clusters cause expression collinearity. Int. J. Dev. Biol. 50, 301-308.
4. Lesne A. 2009. Biologie des systèmes. Medicine/Science 25, 585-587.
5. Izpisua- Belmonte, J.-C., Falkenstein, H., Dollé, P. Renucci, A. & Duboule, D. 1991. Murine genes related to the Drosophila AbdB homeotic gene are sequentially expressed during development of the posterior part of the body. EMBO J. 10, 2279-2289.
6. Dollé, P., Izpisua-Belmonte, J.-C., Brown, J.M., Tickle, C. &Duboule, D. 1991. HOX-4 genes and the morphogenesis of mammalian genitalia. Genes & Dev. 5, 1767-1777.
7. Duboule, D. 2007. The rise and fall of Hox gene clusters. Development 134, 2549-2560.
8. Tarchini, B. & Duboule, D. 2006. Control of Hoxd genes’ collinearity during early limb development. Developmental Cell 10, 93-103.
9. Tschopp. P., Tarchini, B., Spitz, F., Zakany, J. & Duboule, D. 2009. Uncoupling time and space in the collinear regulation of Hox genes. PloS Genetics 5:(3). 10. Morey, C., Da Silva, N.R., Kmita, M., Duboule, D. & Bickmore, W.A. 2008. Ectopic nuclear reorganization driven by a Hoxb1 transgene transposed into Hoxd. J. Cell Sci. 121, 571-577.
11. Tschopp, P. & Duboule, D. 2011. A regulatory ‘landscape effect’ over the Hoxd cluster. Dev. Biol. 351, 288-296.
12. Papageorgiou, S. 2009. A biophysical mechanism may control the collinearity of Hoxd genes during the early phase of limb development. Hum Genomics 3, 275-280.
13. Papageorgiou, S. 2011. Physical forces may cause Hox gene collinearity in the primary and secondary axes of the developing vertebrates. Develop. Growth & Differ. 53, 1-8.
14. Papantonis, A. & Cook, P.R. 2011. Fixing the model for transcription: The DNA moves, not the polymerase. Transcr. 2, 41-44.
15. Bartlett
16. Kondo, T. 1999. Breaking collinearity in the mouse HOXD complex. Cell 97, 407-417.
17. Yamagishi, T., Ozawa, M., Ohtsuka, C., Ohyama-Goto, R. & Kondo, T. 2007. Evx2-Hoxd13 intergenic region restricts enhancer association to Hoxd13 promoter. PLoS ONE Issue 1 (e175).
Spyros Papageorgiou
NCSR ‘Demokritos’, Athens , Greece
Email:spapage@bio.demokritos.gr
------
De profundis – apology and denunciation.
With the opportunity of the above exposition, I would like to put in writing some thoughts about publications, peer reviewing and models.
More often than not, my papers were rejected by editors and referees who advanced frustrating if not erroneous justifications. A common objection brought forward was that my papers do not supply new data. Another disapproval in different versions was that the proposed phenomena and molecules have not been observed before etc.
Such arguments, popular among some biologists, deserve commenting which might prove useful.
1. Because of the increasing complexity in the methodologies involved, practicing scientists in all scientific branches that have come of age are either experimentalists or theorists. For instance and for a long time now, in Physics this is the rule almost without exception. However, the advancement of knowledge is achieved only when Experiment and Theory cooperate tightly. In many cases the activity in one is a challenge for the other: the explanation of an unexpected experimental finding is a challenge for the theoretician. Vica versa, a prediction according to a theory is a challenge for the experimentalist either to demolish the theory or broaden its range of validity. I think that the production of new data on one hand and their correct interpretation on the other are equally important and should be put on equal footing.
In many biological fields (e.g. structural biology, biophysics etc) the cooperation of experiment and theory is quite impressive and fruitful. Unfortunately I cannot maintain that this spirit of teamwork applies in Developmental Biology. Look for instance in the published developmental biology papers: how many refer to some theoretical publication? This occurs very rarely. Is it because there is no activity in this direction? I believe this is not the case. I think the reason is that several developmental biologists consider their work should be completed by an explanatory model which they feel they are self-sufficient to produce. This leads to an estrangement of theorists from their experimental colleagues. There are some laudable incitements for cooperation (e.g. J. Lewis Science 322, 399 (2008)) which indirectly ascertain the absence of collaborative interaction between the two groups. I hope this situation will gradually change. Personally, on several occasions, I have worked harmoniously side by side with my experimental colleagues.
Development has progressed in a spectacular way the last thirty years and I am sure the whole field would benefit much more if the scientists involved were prepared to leave the explanation of their data to other competent professionals. This task is demanding and a satisfactory interpretation of the experiments requires the use of mathematically dressed ideas. Quantitative (mathematical) elaborations should substitute simplistic or qualitative descriptions. An analysis in depth of the data is not a sheer computational problem but it is mainly a conceptual one which requires, in many cases, the application of novel ideas imported from other disciplines.
2. Scientific discoveries are usually the result of unexpected observations in experiments set up by ingenious researchers. However, it is a naïve attitude to believe and trust events and facts that we only observe. If this stand were adopted we would miss many important phenomena in the world. For instance, neutrinos are not and cannot be directly observed but no-one doubts they are abundantly present in the whole Universe moving in all directions (with the speed of light?). We are indirectly sure they exist since this is a necessity based on a First Principle - the energy-and-momentum-conservation axiom that we trust (as yet).
At this point I would like to go a step further and claim that, as long as a phenomenon does not contradict some First Principle, it should be considered seriously as a working hypothesis. At the same time one should anticipate corollary phenomena that could be susceptible of verification. The physical forces proposition of the biophysical model is such a working hypothesis to be further tested.
In conclusion I think that, even if not fashionable and trendy, a well justified assumption aiming at explaining the experimental results should be encouraged by journal editors and referees. Historically even if initially rejected, a right hypothesis is recognized in retrospect sooner or later.
3. I am grateful to the editors and reviewers of the journals who accepted my papers despite a general spirit of mistrust. I am particularly indebted to the editor and referees of Development, Growth & Differentiation for their thoughtful comments and suggestions that helped me improve my manuscript published in this journal in 2011.